The endocrine system 8: kidneys, heart and skin
The endocrine system comprises glands and tissues that produce hormones for regulating and coordinating vital bodily functions. This article, the last in an eight-part series, looks at the endocrine functions of the skin, heart and kidneys
Abstract
The endocrine system comprises glands and tissues that produce hormones to regulate and coordinate vital bodily functions. This article – the last in an eight-part series – discusses the secondary endocrine function of the kidneys, heart and skin.
Citation: Taylor J, Knight J (2021) Endocrine system 8: kidneys, heart and skin. Nursing Times [online]; 117: 12, 46-50.
Authors: James Taylor is lecturer in anatomy and physiology; John Knight is associate professor in biomedical science; both at the College of Human and Health Sciences, Swansea University.
- This article has been double-blind peer reviewed
- Scroll down to read the article or download a print-friendly PDF here (if the PDF fails to fully download please try again using a different browser)
- Click here to see other articles in this series
Introduction
The endocrine system is made up of tissues and glands that produce hormones to coordinate and regulate vital bodily functions. Previous articles in this series have examined the anatomy and physiology of the major dedicated endocrine glands; however, many other organs and tissues have a secondary endocrine function. In this last article of the eight-part series, we explore the endocrinology of the kidneys, heart and skin.
Kidneys
Situated in the upper retroperitoneal region of the abdomen on both sides of the vertebral column, the kidneys are essential components of the wider urinary system, along with the ureters, bladder and urethra. Their primary functions are to:
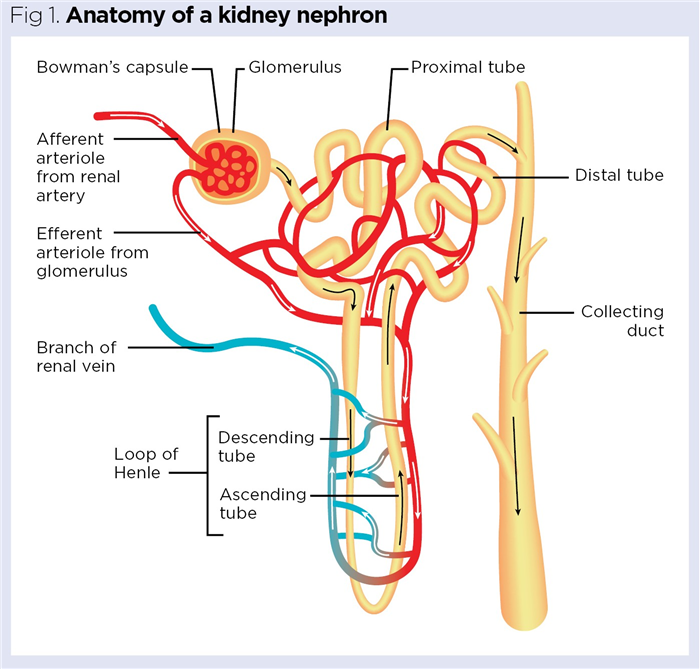
This is facilitated by the nephrons, the structural and functional units of the kidneys (Fig 1). Each kidney contains approximately 1.25 million nephrons (Cook et al, 2021), with healthy adult kidneys filtering approximately 120mL of blood plasma per minute (Gueutin et al, 2012). Closely linked to this role are regulatory roles in:
- Blood volume and blood pressure;
- Blood pH;
- Erythropoiesis (erythrocyte/red blood cell (RBC) production);
- Thrombopoiesis (platelet production) (Gueutin et al, 2012).
Endocrine functions
The kidneys produce a range of hormones with autocrine, paracrine and endocrine effects, which regulate aspects of our wider biology, including erythrocyte and thrombocyte production and calcium ion concentration. Three main hormones produced by the kidneys are:
The kidneys also produce the enzyme renin, an aspartic protease that, although not commonly referred to as a hormone, plays a key endocrine role in blood-pressure regulation via initiation of the renin-angiotensin-aldosterone system (RAAS).
Alongside its role as an endocrine organ, the kidney is itself a key target organ for several hormones produced by other endocrine glands, including those of the heart and adrenal glands.
“The kidneys produce hormones with autocrine, paracrine and endocrine effects”
Erythropoietin
During foetal development, EPO is primarily produced by the liver hepatocytes. Following birth and throughout life, the primary site of production becomes the interstitial peritubular fibroblasts of the renal cortex that surround the nephrons (Jelkmann, 2011). Production and release of the peptide hormone EPO, and expression of EPO receptors (EPO-Rs), is triggered by low oxygen in the tissues (hypoxia), commonly resulting from ischemia, tissue damage, anaemia or high altitude (Tsiftsoglou, 2021).
EPO is the primary hormone regulating erythropoiesis, involving the maturation of erythroid progenitor cells into mature RBCs in the bone marrow (Wenger and Kurtz, 2011). Once released into the bloodstream, EPO binds EPO-Rs on the surface of the RBCs and erythroid progenitor cells, initiating a series of biochemical signaling pathways.
The pathways activated trigger proliferation and alteration of the erythroid progenitor cells which, upon maturation, give rise to further RBCs. Alteration of progenitor cells prevents their removal through apoptosis (programmed cell death). Additionally, EPO binding induces fixed differentiation of the progenitor cells. Both processes ensure progenitor cells mature into RBCs, serving to increase RBC mass (Hedley et al, 2011). By increasing the number of RBCs in the peripheral blood in this way, EPO counters hypoxia by promoting processes that improve tissue oxygenation (Lombardero et al, 2011).
EPO also affects the body in other ways, with complexes of EPO-Rs found in tissues including the heart, brain and skeletal muscle (Tsiftsoglou, 2021). Although the extent of the effects of EPO are not yet fully understood, roles have been noted in:
- Cellular protection;
- Promotion of angiogenesis (formation of new blood vessels);
- Induction of stem-cell maturation pathways, leading to the remodelling of bone (Tsiftsoglou, 2021).
Thrombopoietin
The kidneys are also involved in regulating the number of circulating platelets (thrombocytes). Together with the liver, the kidneys secrete TPO, a 332-amino-acid glycoprotein also known as megakaryocyte growth and development factor. This is produced by cells of the proximal convoluted tubule, before circulating in the plasma and stimulating the resident megakaryocytes of the red bone marrow to enhance their differentiation and fragmentation into platelets. Current evidence suggests the liver is the major producer of TPO, with levels supplemented by the kidneys (Hitchcock et al, 2021).
Calcitriol and activation of vitamin D
Vitamin D, also known as calciferol, represents a group of secosteroids, of which the two primary forms are:
Vitamin D2 is produced by plants, with synthetic versions often used to fortify foods, while D3 is obtained through the action of ultraviolet (UV) light on the skin as well as dietary intake (Ross et al, 2011). These forms of vitamin D are biologically low activity and often considered prohormones (hormone precursor molecules) of the more-active form (calcitriol) with greater hormonal action.
Activation of vitamin D occurs via a two-step metabolic pathway: the liver converts vitamin D into an inactive form called calcidiol; this is then converted into the active form calcitriol in the cells of the proximal convoluted tubule of the kidney (Wang et al, 2015).
Calcitriol, a steroid hormone, is principally responsible for stimulating increased reabsorption of calcium from the intestine, which increases blood calcium (Vieth, 2020). Although the entire intestine is affected by calcitriol action, the primary targets are the duodenum and jejunum (Ross et al, 2011). This reabsorption of calcium, and resultant elevation in the blood, serves to maintain bone structure by providing sufficient calcium for bone mineralisation.
Calcitriol also directly targets the parathyroid glands, regulating the number of calcium receptors present and inhibiting production of parathyroid hormone, thereby preventing bone demineralisation (Hinson et al, 2010).
Renin and the renin-angiotensin-aldosterone system
The kidneys are essential in regulating blood pressure, both by controlling plasma volume (by regulating the volume of urine produced) and renin production to initiate the RAAS. (See part 4 of this series for full details of the adrenal glands.)
Heart
The heart forms the core of the circulatory system, pumping deoxygenated blood to the lungs and oxygenated blood throughout the body. Structurally, it comprises three main layers:
- Pericardium (outermost covering) – protects the heart from overexpansion and infection;
- Myocardium or cardiac muscle (central layer) – contains cardiomyocytes responsible for the heart’s contractile nature and most of its endocrine function;
- Endocardium – lines the inner structures of the heart.
Endocrine function
Alongside its primary function of blood circulation, the heart works as a secondary endocrine organ, producing hormones that act, principally, on the kidneys to help regulate blood–sodium concentration, extracellular volume and blood pressure (Lugnier et al, 2019; Ogawa and de Bold, 2014). Two of these endocrine primary hormones are:
Atrial and brain natriuretic peptides
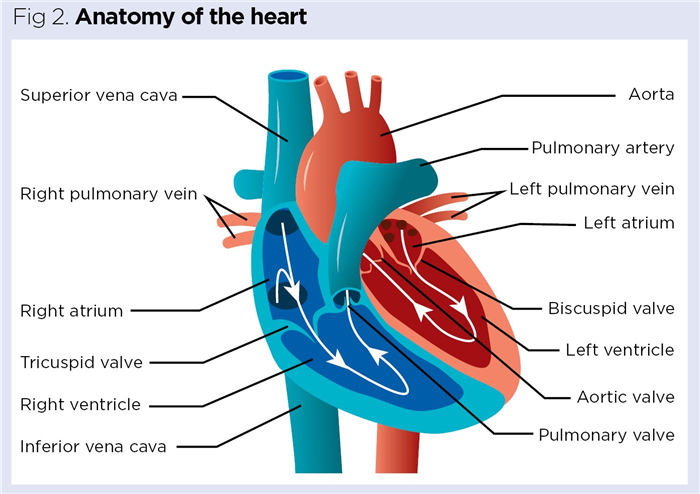
Central to the heart’s primary function as a pump are four chambers: left and right atrium; and left and right ventricles (Fig 2). ANP (also called atrial natriuretic factor and A-type natriuretic peptide) and BNP (also called B-type natriuretic peptide) are primarily secreted from the atria. Lower levels of ANP and BNP are also secreted from the ventricles and aortic arch of the heart, as well as other tissues, including those of the pituitary gland, hypothalamus and lungs (Ogawa and de Bold, 2014).
Released into the blood in response to the stretching of the heart’s myocardium, both ANP and BNP are secreted by the cardiomyocytes and induce several similar effects (Nakagawa et al, 2019). As natriuretic peptides, their principal effects are to promote natriuresis, acting on the kidneys to increase the excretion of sodium in the urine. By decreasing blood–sodium concentration, they also cause a reduction in blood volume and, therefore, blood pressure.
The heart’s hormonal influence on kidney function, along with its intrinsic effects on cardiovascular function, means it is often considered to be self-regulating (Ogawa and de Bold, 2014).
Three main forms of natriuretic peptide receptor (NPR) have been identified:
Of these, NPR-A is the preferential receptor for ANP and BNP binding, shown to be expressed throughout several tissue types, including those of the kidneys, adrenal glands, brain and heart (Potter et al, 2009). For both ANP and BNP, NPR-C acts as a clearance receptor which, on binding the active forms of the natriuretic peptides, removes and inactivates them, thereby regulating their effects.
Once released into the blood, ANP and BNP induce two key effects in the heart:
Both increase the overall elasticity of the heart, facilitating a decrease in blood pressure (Nakagawa et al, 2019). Similarly, both natriuretic peptides inhibit the secretion of aldosterone from the adrenal cortex and renin from the kidney – two elements of the RAAS serving to increase blood pressure. The net effects of natriuretic peptide release are, therefore, in direct contrast with those of the RAAS (Nakagawa et al, 2019).
Aside from their inhibition of renin secretion, ANP and BNP have natriuretic and diuretic effects on the kidneys through their targeting of the nephrons. First, they increase the glomerular filtration rate (GFR) by causing vasodilation of the afferent arteriole (Fig 1), thereby increasing the volume of blood entering the glomerulus for filtration (Wong et al, 2017). They also act on the intraglomerular mesangial cells (found between the glomerular capillaries), causing their relaxation and allowing expansion of the capillaries themselves; this greater surface area, again, increases filtration (Wong et al, 2017; Theilig and Wu, 2015). Secondly, the natriuretic peptides target the collecting duct (Fig 1), preventing reabsorption and increasing excretion of sodium. The collecting duct is the final site of the nephron from which sodium can be reabsorbed.
Prevention of sodium reabsorption is achieved via two main actions, each targeting the principal means by which sodium is transported in the collecting duct. The collecting duct contains:
- Apical sodium channels for passive transport (requiring no energy);
- Basolateral sodium-potassium adenosine triphosphatase (ATPase) pumps for active transport (requiring energy).
Natriuretic peptides induce the inhibition of these sodium transport mechanisms through the cyclic nucleotide secondary messenger molecule, cyclic guanosine monophosphate (cGMP). cGMP-mediated inactivation of these transport mechanisms allows for increased sodium excretion from the body by preventing its reabsorption (Wong et al, 2017). In this way, water remains in the tubules and is removed with sodium in urine. The net effects of natriuretic peptides are, therefore, to increase urine volume and decrease blood volume and pressure, by increasing the excretion of sodium from the body.
Skin
Alongside the hair, nails and associated exocrine glands, the skin is a core element of the integumentary system, which is the largest organ in the human body. It has several key functions that are crucial to the homeostasis of multiple aspects of our biology. Four primary functions are:
- Protection from damage (for example, from radiation, infection, or physical, chemical and thermal influences);
- Regulation of body temperature;
- Sensation of the external environment through nerve endings;
- Synthesis of vitamin D (Abdo et al, 2020).
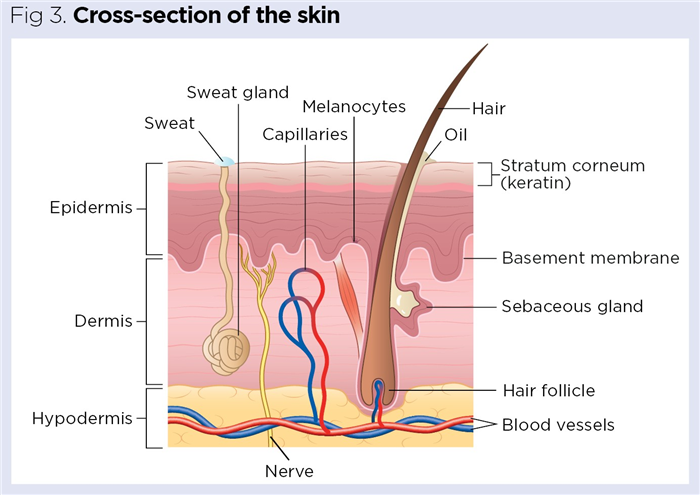
At the core of the skin’s key functions is its structure, which comprises three main layers (Fig 3):
- Epidermis – protective waterproof outer covering, composed of stratified squamous epithelium;
- Dermis – predominantly connective tissue, giving structure and elasticity, but also contains blood vessels, nerves and exocrine glands for the production of sweat (via the sweat glands) and sebum (via the sebaceous glands);
- Hypodermis – largely connective tissue (adipose or areolar) with an extensive network of blood vessels. Its adipose composition serves as both energy reserve and insulation (Cook et al, 2021).
Endocrine function
Although the skin is often overlooked as an endocrine organ, it is involved both in the regulation (through activation, inactivation and removal) and the production of hormones. Hormones produced include:
- Insulin-like growth factor-1 (IGF-1);
- Steroid hormones including sex hormones (androgens and oestrogens);
- Melanocyte-stimulating hormone (MSH);
- Vitamin D (Farag et al, 2019; Zouboulis, 2009).
Insulin-like growth factor-1
As mentioned in part 6 of this series (which explores the endocrinology of the pancreas, stomach, small intestine and liver), IGF-1 (formerly known as somatomedin) is a polypeptide hormone that is structurally similar to insulin. It is primarily produced by the hepatocytes of the liver and its receptors are present in numerous body tissues (Taylor and Knight, 2021). Secreted in response to growth hormone released from the anterior region of the pituitary gland, upon binding its receptor, IGF-1 initiates several secondary messenger signalling pathways to bring about growth and tissue maturation (Kineman et al, 2018).
In the skin, IGF-1 is produced in the dermis from cells called dermal fibroblasts (Farag et al, 2019). IGF-1 has widespread effects on cellular growth and development, particularly during puberty, when levels rise sharply (Cole et al, 2015).
IGF-1 also has several effects on the skin and wider integumentary system; key among these are effects on the:
IGF-1 affects the hair growth cycle as well as the proliferation and differentiation of follicles – crucial to growth and maintenance. It is also involved in the remodelling of tissue (Trüeb, 2018).
During the epidermis’ response to UVB exposure, IGF-1 plays a key role in facilitating an appropriate keratinocyte response, preventing the possible malignant transformation of cells. The epidermis, comprised principally of keratinocytes, contains numerous IGF-1 receptors that are activated by IGF-1 in response to UVB-induced cellular damage. When receptors are not activated, this can lead to a proliferative response and, potentially, skin cancer; this is particularly important in older people as levels of IGF-1 expression decrease with age (Lewis et al, 2010).
Steroid hormone production
An often overlooked, and more recently established endocrine role of the skin, is in the production and metabolism of steroid hormones. As well as expressing several steroid-hormone receptors, skin cells possess all the biochemical elements required for the production of hormones, including androgens, oestrogens and glucocorticoids (Slominski et al, 2013). Notable hormones produced include:
- Cortisol;
- Corticosterone;
- Some of the main sex hormones – namely, oestradiol (the primary form of oestrogen), dihydrotestosterone and testosterone.
Local production of these hormones is typically regulated by the localised production of adrenocorticotropic hormone and corticotropin-releasing hormone by the skin (Slominski et al, 2013). In addition, glucocorticoid production in the skin is increased in response to UVB exposure (Skobowiat et al, 2013). In the skin, oestradiol and testosterone maintain the integrity of the extracellular matrix by inducing collagen production, which is essential to the overall elasticity and structure of the dermis (Haag et al, 2012). Levels of testosterone (and its precursor, dehydroepiandrosterone) decrease with age; this contributes to wrinkles and the loss of elasticity – two visible signs of skin ageing (Haag et al, 2012).
“Often overlooked, and one of the more recently established elements of the skin as an endocrine organ, is its role in the production and metabolism of steroid hormones”
Melanocyte-stimulating hormone
The epidermis contains cellular arrangements, known as epidermal units, each of which comprises a single melanocyte surrounded by 36 keratinocytes (Maranduca et al, 2019). Through paracrine regulation between both cell types, the melanocytes are responsible for skin pigmentation and protection against the effects of UV light through the production of melanin.
Melanogenesis is triggered by the release of MSH (also called melanotropin), produced primarily by the intermediate lobe of the pituitary gland, but also by the skin (Videira et al, 2013). MSH represents a group of peptide hormones derived from the peptide precursor, pro-opiomelanocortin (POMC), a key peptide in the melanocortin system responsible for regulating food intake and energy expenditure (Hill and Faulkner, 2017; Nuzzaci et al, 2015). POMC gives rise to several biologically active regulatory peptides, called melanocortins, of which MSH is one group.
In response to UV light, melanocytes secrete MSH. On binding to melanocortin receptors on the surface of melanocytes, the production and release of melanin increases; it is stored in structures called melanosomes (Maranduca et al, 2019). Melanin is also redistributed in cells, with melanosomes transferred into the surrounding keratinocytes making up each epidermal unit. Melanin, therefore, provides a crucial role in the homeostasis of the skin: through its ability to absorb and/or reflect UV light, it has a photoprotective role in limiting cellular damage from UV exposure (Maranduca et al, 2019).
Vitamin D
As mentioned earlier, vitamin D represents a group of secosteroids, of which the primary form, vitamin D3 (cholecalciferol), is principally synthesised through the action of UV light on the skin. When skin is exposed to UVB, it causes photolysis of the steroid, 7-dehydrocholesterol (provitamin D3) forming previtamin D3; which, after thermal isomerisation, gives rise to vitamin D3 (Bikle, 2012).
Being relatively inactive – and, therefore, generally considered a prohormone – vitamin D3 is transported via the blood from the skin to the liver, bound by a vitamin D-binding protein. Once in the liver, vitamin D3 is converted to calcidiol (25-hydroxyvitamin D3), the main form of vitamin D found in circulation (Bikle, 2012). Following transport to the kidneys, calcidiol is then further converted in the cells of the proximal convoluted tubules, producing the active form of vitamin D, calcitriol (1α,25-dihydroxyvitamin D3); this is responsible for the majority of hormonal effects discussed (Wang et al, 2015).
Conclusion
The endocrine system is a complex series of glands and tissues that produce and secrete hormones, which are used to regulate and coordinate vital bodily functions. This article on the secondary endocrine functions of the kidney, heart and skin concludes our series in which we explored each of the major endocrine glands and their hormonal secretions.
Key points
- The kidneys, heart and skin have secondary endocrine functions
- A key hormone secreted by the kidneys is erythropoietin, which counters hypoxia
- Hormones produced by the heart act principally on the kidneys, helping to regulate blood-sodium, extracellular volume and blood pressure
- The steroid hormone calcitriol helps maintain bone structure and is produced by the kidneys through the activation of vitamin D
- The skin is often overlooked as an endocrine organ, but regulates and produces various hormones, along with the prohormone vitamin D3
Also in this series
Abdo JM et al (2020) The applied anatomy of human skin: a model for regeneration. Wound Medicine; 28: 100179.
Bikle DD (2012) Vitamin D and bone. Current Osteoporosis Reports; 10: 2, 151-159.
Cole TJ et al (2015) The relationship between Insulin-like Growth Factor 1, sex steroids and timing of the pubertal growth spurt. Clinical Endocrinology; 82: 6, 862-869.
Cook N et al (2021) Essentials of Anatomy and Physiology for Nursing Practice. SAGE.
Farag AGA et al (2019) Role of insulin-like growth factor-1 in skin tags: a clinical, genetic and immunohistochemical study in a sample of Egyptian patients. Clinical, Cosmetic and Investigational Dermatology; 12: 255-266.
Gueutin V et al (2012) [Renal physiology]. Bulletin du Cancer; 99: 3, 237-249.
Haag M et al (2012) Age and skin site related differences in steroid metabolism in male skin point to a key role of sebocytes in cutaneous hormone metabolism. Dermato-endocrinology; 4: 1, 58-64.
Hedley BD et al (2011) The role of erythropoietin and erythropoiesis-stimulating agents in tumor progression. Clinical Cancer Research; 17: 20, 6373-6380.
Hill JW, Faulkner LD (2017) The role of the melanocortin system in metabolic disease: new developments and advances. Neuroendocrinology; 104: 4, 330-346.
Hinson J et al (2010) The Endocrine System. Churchill Livingstone.
Hitchcock IS et al (2021) The thrombopoietin receptor: revisiting the master regulator of platelet production. Platelets; 32: 6, 770-778.
Jelkmann W (2011) Regulation of erythropoietin production. Journal of Physiology; 589: Pt 6, 1251-1258.
Kineman RD et al (2018) 40 years of IGF1: understanding the tissue-specific roles of IGF1/IGF1R in regulating metabolism using the Cre/loxP system. Journal of Molecular Endocrinology; 61: 1, T187-T198.
Lewis DA et al (2010) The IGF-1/IGF-1R signaling axis in the skin: a new role for the dermis in aging-associated skin cancer. Oncogene; 29: 10, 1475–1485.
Lombardero M et al (2011) Erythropoietin: a hormone with multiple functions. Pathobiology; 78: 1, 41–53.
Lugnier C et al (2019) The endocrine function of the heart: physiology and involvements of natriuretic peptides and cyclic nucleotide phosphodiesterases in heart failure. Journal of Clinical Medicine; 8: 10, 1746.
Maranduca MA et al (2019) Synthesis and physiological implications of melanic pigments. Oncology Letters; 17: 5, 4183-4187.
Nakagawa Y et al (2019) Atrial and brain natriuretic peptides: hormones secreted from the heart. Peptides; 111: 18-25.
Nuzzaci D et al (2015) Plasticity of the melanocortin system: determinants and possible consequences on food intake. Frontiers in Endocrinology; 6: 143.
Ogawa T, de Bold AJ (2014) The heart as an endocrine organ. Endocrine Connections; 3: 2, R31-R44.
Potter LR et al (2009) Natriuretic Peptides: their structures, receptors, physiologic functions and therapeutic applications. In: Schmidt HHHW et al (eds) cGMP: Generators, Effectors and Therapeutic Implications. Handbook of Experimental Pharmacology, Vol 191. Springer.
Ross AC et al (2011) Dietary Reference Intakes for Calcium and Vitamin D. The National Academies Press.
Sahay M et al (2012) Renal endocrinology: the new frontier. Indian Journal of Endocrinology and Metabolism; 16: 2, 154-155.
Skobowiat C et al (2013) Ultraviolet radiation regulates cortisol activity in a waveband dependent manner in human skin ex-vivo. British Journal of Dermatology; 168: 3, 595-601.
Slominski A et al (2013) Steroidogenesis in the skin: implications for local immune functions. The Journal of Steroid Biochemistry and Molecular Biology; 137: 107-123.
Taylor J, Knight J (2021) Endocrine system 6: pancreas, stomach, small intestine and liver. Nursing Times [online]; 117: 10, 46–50.
Theilig F, Wu Q (2015) ANP-induced signaling cascade and its implications in renal pathophysiology. American Journal of Physiology: Renal Physiology; 308: 10, F1047-F1055.
Trüeb RM (2018) Further clinical evidence for the effect of IGF-1 on hair growth and alopecia. Skin Appendage Disorders; 4: 2, 90-95.
Tsiftsoglou AS (2021) Erythropoietin (EPO) as a key regulator of erythropoiesis, bone remodeling and endothelial transdifferentiation of multipotent mesenchymal stem cells (MSCs): implications in regenerative medicine. Cells; 10: 8, 2140.
Videira IF et al (2013) [Mechanisms regulating melanogenesis]. Anais Brasileiros de Dermatologia; 88: 1, 76-83.
Vieth R (2020) Vitamin D supplementation: cholecalciferol, calcifediol, and calcitriol. European Journal of Clinical Nutrition; 74: 1493-1497.
Wang Y et al (2015) The vitamin D receptor in the proximal renal tubule is a key regulator of serum 1α,25-dihydroxyvitamin D3. American Journal of Physiology Endocrinology and Metabolism; 308, E201–E205.
Wenger RH, Kurtz A (2011) Erythropoietin. Comprehensive Physiology; 1: 4, 1759-1794.
Wong PC et al (2017) The renal and cardiovascular effects of natriuretic peptides. Advances in Physiology Education; 41: 2, 179-185.
Zouboulis CC (2009) The skin as an endocrine organ. Dermato-endocrinology; 1: 5, 250-252.